

蜘蛛是洞穴生态系统中最主要的捕食者之一,有超过1000种物种专门在地下发现,其相同数量的物种在地下栖息地度过了生命周期的一部分。Orb蜘蛛是研究行为适应的优秀模式生物,由于它们高度结构化的二维网构成了其网构建行为的直接记录,所以也是觅食行为,可以通过测量照片中的大量几何变量轻松量化。
标准的球形网由一个轮毂组成,蜘蛛通常居住在那里,放置在椭圆形网的几何中心上方,半径从中向外辐射,就像轮子上的辐条一样,一个粘性的捕获螺旋覆盖半径,整个结构被封闭在一个框架内,许多系泊线从框架中将网连接到周围的植被。

球体腹板几何形状
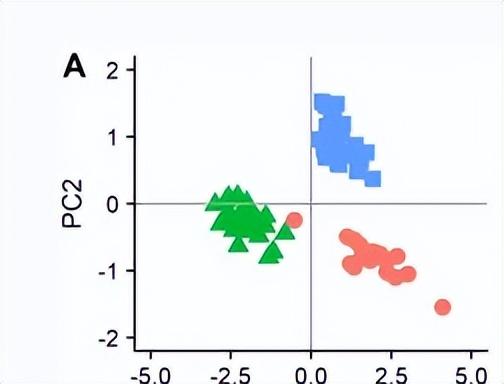
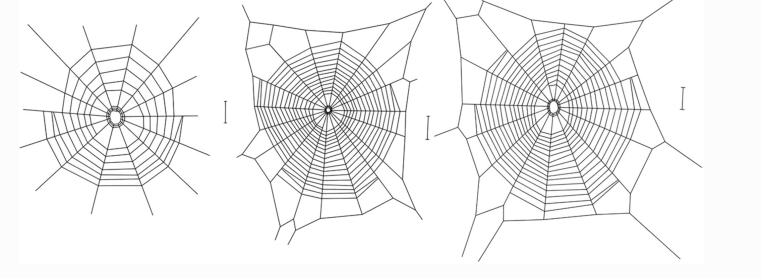
对这三个物种的网的整体结构进行视觉比较,强烈表明它们之间的差异,洞穴蜘蛛M. menardi的网较小,螺旋较少,最明显的是框架线很少或没有框架线。单独分析网状时,发现相对腹网面积(总网面积除以头胸宽度平方)存在显著差异,其中梅纳尔迪分枝杆菌网小于两种地上物种的网。
我们观察到穴居球状蜘蛛Meta menardi的网,与相关地上物种的网在统计学上可辨别和视觉上明显的差异,最值得注意的是缺少通常包围球形腹板的捕获螺旋和半径的框架。这导致半径通过直接附着在周围的岩石基质上来充当系泊螺纹。
这些洞穴网比标准球网具有更多的系泊线,这可以向蜘蛛提供有关猎物在网附近岩石表面上行走的信息。据我们所知,这项研究利用蜘蛛网提供的独特机会来量化觅食行为,是极少数关于穴居动物的研究之一,与密切相关的地表动物相比,这些动物的行为存在显着差异。
对地下环境的形态适应的研究比行为适应要详细得多,在许多特征中都有趋同进化的例子,包括眼睛的丧失,色素的丧失和肢体伸长。我们比较了洞穴球蜘蛛的肢体伸长,因为它们没有显示出眼睛或色素损失的证据。

然而,与我们的假设相反,M. menardi在I或III腿上都没有最长的相对腿长,尽管它的相对腿长确实比其最近的表面亲戚Metellina mengei具有统计学上的显着长,兼性洞穴蜘蛛Metellina merianae在这两个物种之间具有相对的腿长,所以需要更详细的研究来确定洞穴球蜘蛛中是否存在一定程度的肢体伸长。
M. menardi的大整体尺寸也可能被视为对地下环境的适应,因为体型增加是洞穴蛛形纲动物中常见的形态适应,蜘蛛通常是四颌蜘蛛科中最大的成员之一,它们的大小可能表明某种程度的形态适应,尽管值得注意的是,大型地表栖蜘蛛在热带地区相对常见。
在洞穴球蜘蛛Meta japonica中,观察到与我们发现的M. menardi非常相似的网状结构,显示出完整框架的遗漏(框架截面数,平均±SD:1.1±0.6)和直接附着在墙上的相似数量的半径(14.6±5.3),然而,捕获螺旋转弯的次数(17.0±4.3)大大大于本研究中梅纳尔迪分枝杆菌的记录(6.0±1.7),这表明网格高度会更低。
洞穴球蜘蛛的饮食行为
从几项实地研究中证实,M. menardi的饮食由36%至69%的非飞行猎物组成,包括蛞蝓,蜘蛛和千足虫,与在其他球状蜘蛛中观察到的主要由飞行猎物组成的饮食相比,人们普遍认为,Meta蜘蛛将传统的网内猎物捕获与新颖的网外狩猎相结合,以实现其扩大的饮食。
然而,涉及独立于网络的狩猎的觅食行为似乎不太可能,因为这需要进化出一种全新的额外猎物捕获行为。相反,半径附着在墙上,M. menardi位于其网的中心,这似乎是它的通常位置,它将能够通过通过半径传递的相同振动感觉机制来检测飞入网中的猎物和行走的猎物与半径碰撞的存在和方向。
在猎物与半径相撞后,M. menardi “迅速将猎物从网追踪到墙上”,并且由Peter Smithers进行的一项未发表的试点研究表明,实验室中的梅纳尔迪 M. menardi 在木虱走进放射状线后,通过跑下来并将丝线连接到木虱上来捕捉木虱。

从这项研究中观察到的洞穴球蜘蛛网中缺乏框架线很可能是一种行为适应,以扩大饮食范围,以应对地下栖息地飞行猎物的稀有性。为了证实这一假设,需要仔细的实地工作或实验室实验,导致直接观察从墙上捕获的猎物,并结合对已确认的地表栖Meta物种之一的网结构和猎物捕获的比较分析。
该比较方法有些偏差,导致所有测量的洞穴蜘蛛都是晚龄幼体,而测量的地上蜘蛛都是成年蜘蛛。虽然这可能会影响我们的一些定量结果,但它不太可能对本文提出的定性差异和结论产生任何影响,因为球状网通常不会发生强烈的个体发生变化。

它们确实显示出的变化与网的不对称性有关,因为较老和较大的蜘蛛由于更快的重力辅助向下运行而构建具有较大下部的网,或者与衍生的网状特征有关,例如幼年蜘蛛中不存在的自由扇区。
这里研究的四颌蜘蛛,除了亚成年洞穴蜘蛛缺乏框架线外,都构建了标准的球状网,并且由于它们构建了倾斜的网,预计大小和不对称之间的联系会很弱。洞穴蜘蛛和本研究中发现的两种地上蜘蛛之间的主要差异。

洞穴球蜘蛛网状结构的变化
在较冷的温度下,球状蜘蛛会形成较小的网,捕获螺旋转弯较少,但半径数量相似,这与我们的发现相似。然而,与我们的发现相反,网孔高度也被发现增加,更重要的是,由于温度,整体卷材结构没有变化。同样,尽管在本研究中没有直接测量,但洞穴中的风可能低于树林和沿河。
较高的风会导致较小的网,捕获螺旋转弯更少,在那里我们发现相反的,在庇护洞穴中最小的网。值得注意的是,对M. mengei的实地研究没有发现,在裸露的林地边缘建造的网和在木材遮蔽的内部建造的网之间在网页设计上有任何差异,同样,风不会影响网络的基本结构。

Meta japonica中框架线的减少是由于在洞穴墙壁和天花板上的小空腔中构建网的结果,正如M. menardi也观察到的那样。行为灵活性似乎发挥了一些作用,因为当框架螺纹存在时,它们主要存在于离岩壁较远的地方,这可能表明框架的存在取决于环境的拓扑结构。
然而,在实验室空间限制下的球状蜘蛛试图通过拉长它们的网以匹配可用空间和降低网眼高度来维持整体网面积,这与我们对M. menardi的发现相反。大多数面临空间限制的物种不会改变其网的结构,尽管已经观察到Araneid Eustala illicita和四颌形Leucauge argyra在非常受限的人造框架和管中构建球状网时将半径直接附着在表面上。
然而,这些腹板只缺少一小部分框架螺纹,这与我们几乎完全消除框架螺纹的发现相反。Anapid Anapisona simoni在框架,径向和捕获线程方面显示出更显着的变化,与本研究中观察到的反应有些相似。
但蹼蹼与四颌蹼有很大不同,因为它们更立体,缺乏不粘性的脚手架螺旋,总体而言,实验室中强烈约束的蜘蛛,和野外的洞穴蜘蛛之间的比较仅具有有限的有效性,因为天然洞穴通常提供一系列潜在的网站,如同源Metellina merianae的正常洞穴球网所证明的那样。

观测到的变化也可能是对地下生境中食物供应不足的直接反应,虽然已知球形蜘蛛会因饥饿而改变生活史和丝绸特性,仅发现球体几何形状的微小定量变化,包括面积增加,但捕获螺旋数量减少,与本研究中观察到的大规模定量和定性差异相反。
梅塔梅纳尔迪没有表现出对饥饿的任何特定的生理适应,总体而言,我们在M. menardi网中观察到的变化不太可能仅由行为灵活性引起,尽管需要对洞穴蜘蛛的网构建行为进行更详细的实验室研究以完全消除这种可能性。

数据收集:网页几何
位于田间的所有网都用水雾化,以使丝线更明显,因此更容易测量。五个不同长度的字符。使用数字游标卡尺测量到最接近毫米:(1)垂直(dv) 和 (2) 水平直径 (dh) 从最外层和相对的螺旋螺纹 (3) 上部 (ru) 和 (4) 水平半径 (rr)从中心轮毂的中心到最外层的螺旋螺纹进行测量。
典型的四颌球网示意图,突出显示了研究中测量的变量。(A)显示不同的丝线类型以及网如何分为4个象限。(B) 显示测量变量:垂直 (dv) 和水平 (dh) 直径,上部 (Ru) 和更低的半径 (rl)和轮毂直径 (H)。

计数测量(图)5B)包括半径数,在4个象限中心(s问1,s问2,s问3,s问4)、框架螺纹的数量和系泊点的数量(即连接腹板和周围基材的系泊螺纹的数量)。
测量的卷材参数使我们能够得出这些进一步的卷材特征:Web 面积(捕获螺旋的面积减去自由扇区和集线器)使用椭圆-集线器公式76 进行估算:(dv2)(dh2)π−(H2)2π较低的垂直半径(rl) 和左水平半径 (rle)的计算方法如下:dv−ru=rldh−rr=rle卷材不对称的特征在于 − 1 到 1 之间的值,0 表示完全垂直对称的卷材。

负值表示轮毂位于比卷材底部更靠近顶部的位置,并使用以下公式找到:ru−rlru+rlWeb形状的特征还在于介于-1到1之间的值,0表示圆形,负值表示它的高度大于宽度,并且使用此公式找到77:dh−dvdh+dv平均网格高度,即两个相邻捕获螺旋转弯之间的距离,是使用此公式测量的。
数据分析:形态学
为了确定梅纳尔分枝杆菌与其表面亲属之间的形态差异(腿I和腿III的总长度,头胸宽度和髌骨 - 胫骨长度),我们使用R中的prcomp函数进行了主成分分析,并使用ggplot2包以生成受戒图。

为了研究梅纳尔迪分枝杆菌的潜在肢体伸长,我们遵循了Hesselberg和Simonsen在线性混合模型中,使用相对腿长(腿I和III的髌骨-胫骨长度除以头胸宽度)作为响应变量来控制蜘蛛的大小,其中物种是固定因子,位置(样带数和洞穴ID)作为随机因子。
确保正态性,所有物种比较中腿I的相对腿长用自然对数变换,为了分离系统发育和栖息地,我们发现与肯沃德-罗杰自由度和Tukey方法的成对对比,用于使用emmeans函数从emmans包中调整P值79作为事后测试,完整模型揭示了物种之间的显着差异。

所有模型都是使用 lme4 包中的 lmer() 函数构建的80在 R 中81.用II型Wald F检验发现P值。数据分析:卷筒纸几何。采用上述方法,建立了半径数、绝对腹板面积、系泊螺纹数、腹板不对称性、腹板形状、相对平均网高和相对腹板面积的线性混合模型,其中物种为固定因子,位置为随机因子。
为了确保正态性,对相对卷材面积进行了对数变换。我们使用上述相同的事后检验进行成对物种比较,其中完整模型显示物种是一个显着变量。所有模型都是使用 lme4 包中的 lmer函数构建的80在 R 中81.用II型Wald F检验发现P值。

我们发现,与两种相关的地表栖球状蜘蛛相比,洞穴球蜘蛛的网状几何形状显示出显着差异,特别是在半径直接附着在洞穴墙壁和天花板上的框架线减少方面,表明这种行为改变可能是为了应对飞行猎物的有限可用性和地下栖息地中行走猎物的高相对丰度。
虽然需要对洞穴球蜘蛛的猎物捕获和建网行为进行更多的观察和实验,但据我们所知,这是第一个提出陆生洞穴节肢动物新行为适应的定量研,这具有更广泛的意义,因为我们研究中的洞穴蜘蛛仅显示出有限的形态适应证据(没有失去眼睛或色素,但与它们最近的表面居住亲戚相比,有一些轻微的肢体伸长的迹象)。

这意味着这些蜘蛛可能处于完全适应洞穴环境的初始阶段,在形态适应之前进行行为适应(最终可能演变成完全失去网和适应网外运动和猎物捕获)。因此,我们认为Meta洞穴蜘蛛可能是研究能量贫乏环境中行为灵活性和行为进化的潜在模式生物。